|
 L'autore: Nino Mannina L'autore: Nino Mannina
PARTE I^
1. GENERALITA'
a. Premessa
Prima di inoltrami nel tema di questo scritto, ci tengo a precisare che a riguardo dello studio del Genere Russula non voglio certo insegnare niente a nessuno, né ho in serbo novità capaci di sconvolgere gli inquadramenti Sistematici ad esso relativi, o roba simile. Sono talmente tanti e di così altissimo livello gli studiosi che si sono occupati e che attualmente si occupano dell’argomento, che il sottoscritto, da quell’illustre sconosciuto che sa perfettamente di essere, non ha nessuna pretesa di aggiungere altro.
Voglio sottolineare quindi, che questo elaborato si indirizza espressamente a coloro i quali vogliono muovere i primi passi verso un esame più accurato di questo peculiare raggruppamento di funghi superiori, il cui studio non è certo privo di difficoltà.
Tuttavia, difficile, di sicuro non significa impossibile.
Tanti appassionati di micologia, infatti si rifiutano categoricamente di avere a che fare con questi macromiceti, ritenendoli talmente ostici, da rinunciare, a priori, ad un qualsiasi approccio determinativo che li riguardi, perdendo, a mio modestissimo avviso, un‘occasione di conoscenza e di approfondimento, sicuramente di grande gratificazione.
Mi propongo allora, attraverso queste righe, di mettere a disposizione la mia modesta esperienza in fatto di Russule, cercando di chiarire quello che dirò, attraverso l’ausilio di disegni esplicativi, tentando così, di rendere comprensibile quello che per sua intrinseca natura lo è poco.
Nutro la sincera speranza, grazie alle pagine di questa sorta di vademecum micologico, di fornire le basi, a chiunque lo volesse, per cominciare a districarsi nell’affascinante universo “Russula”, senza scappare immediatamente altrove, nel caso di un più che probabile incontro, con gli sporofori delle specie appartenenti a questo complesso Genere.
b. Posizione Tassonomica
Le Russule appartengono al Regno dei Funghi (Regno Fungi), che racchiude organismi Eucarioti (cellule contenenti un nucleo delimitato da una membrana), eterotrofi (capaci cioè di nutrirsi attingendo unicamente a sostanza organica preesistente), formati da un tallo vegetativo, detto micelio, costituito da elementi filamentosi tubiformi dette ife, le cui pareti sono costituite da micosina (composto molecolare simile alla chitina).
Sono esseri viventi che non differenziano aggregazioni tissutali aventi precise funzioni di organi. Accumulano nelle cellule, quali riserva energetica, uno zucchero complesso, chiamatoGlicogeno.
Le ife miceliari possono essere settate, ove la delimitazione imposta da due setti opposti circoscrive una singola cellula (articolo ifale), dando luogo quindi a sottilissime catenule di lunghezza indefinita, nelle quali ciascuna di esse comunica con la successiva attraverso un orifizio (doliporo). In tale modo, i contenuti citpoplasmatici possano circolare liberamente lungo il lume interno delle ife stesse. Nel loro insieme, gli ammassi ifali, fondano un organismo pluricellulare (micelio eucarpico), capace di espandersi indefinitamente ed in qualsiasi direzione nel substrato (fonte da cui il micelio trae sostentamento).
Oppure altresì, dare luogo a sistemi ifali del tutto privi di setti, originanti un tallo monocellulare e polinucleato (micelio cenocitico).
In generale, hanno riproduzione agamica oppure sessuata, tramite la differenziazione di spore esogene o endogene rispetto alla cellula sporigena che le ha prodotte (basidi nel primo caso, aschi nel secondo).
Hanno tipologia nutritiva saprotrofa, parassitaria o micorrizogena (simbiotica). Idealmente, fanno parte dei funghi cosiddetti superiori, ovvero capaci di esprimere strutture riproduttive visibili ad occhio nudo (tecnicamente noti come sporofori ovvero portatori di spore, oppure carposomi ovvero con aspetto di frutto, oppure basidiomi o corpi fruttiferi estrinsecati dai basidiomiceti, oppure carpofori, designazione ormai desueta che significa portatori di frutto).
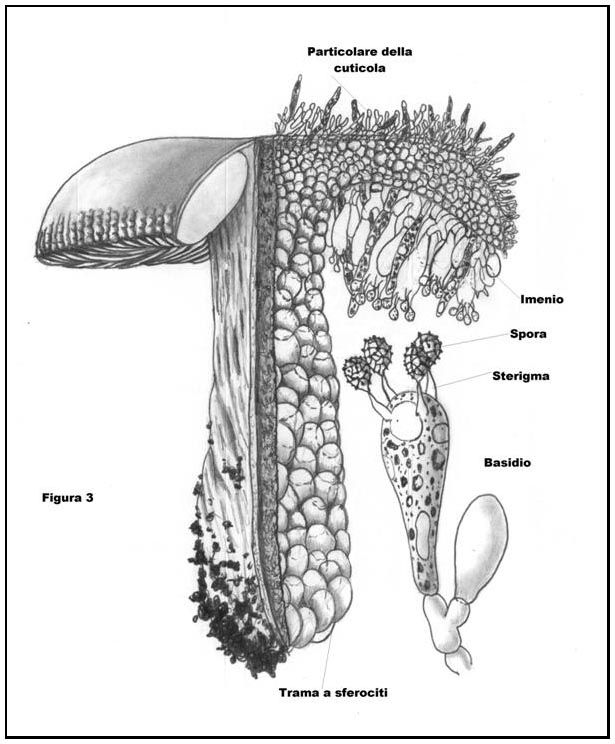
Le Russule, appartengono alla Divisione Basidiomycota e alla Classe Basidiomycetes, hanno cioè sporofori con imenio (che è la parte riproduttiva, in senso stretto, dello sporoforo stesso) costituito da basidi, nei quali le spore (le vere cellule riproduttive dei funghi) maturano al loro esterno, su appendici chiamate sterigmi (cfr. Fig. 3 struttura della trama di una Russula ideale).
L’imenio, nella sua globalità, formato come anzidetto da cellule sporigene chiamate basidi, a cui si mescolano elementi imeniali sterili, quali cistidi e basidioli, è alloggiato sulle facce delle lamelle, costituenti nel loro insieme l‘imenoforo (cioè portatore dell’imenio).
Sono incluse nelle Sottoclasse Holobasidiomycetidae, ovvero sono provvisti di olobasidi (basidi unicellulari).
Fanno parte dell’Ordine Russulales e della FamigliaRussulaceae, nel quale confluiscono sia il Genere Russula che l’affine Genere Lactarius, entrambi accomunati dall’istologica sporoforale, dalle spore con ornamentazioni formate di escrescenze amiloidi, tuttavia difformi per l’emissione di latice alla frattura, assente nelle Russula e ben presente invece nei Lactarius.
Alla sopra citata Famiglia delle Russulaceae appartengono anche i Generi Arcangeliella, Boidinia, Cystangium,Gloecystidiellum, Gymnomyces, Macowanites,Zelleromyces. Riassumendo, l’inquadramento tassonomico delle Russule è il seguente:
Regno: Fungi;
Divisione: Basidiomycota;
Classe: Basidiomycetes;
Sottoclasse: Holobasidiomycetidae;
Ordine: Russulales;
Famiglia: Russulaceae
Genere: Russula.c. Morfologia generale degli sporofori e loro dimensioni
Quel che prima di tutto si nota incontrando qualsiasi sporoforo fungino, sia nei boschi che in qualunque altro habitat capiti di osservarne la presenza, sono le forme che questi possiedono. Ed ogni identificazione sul campo, passa necessariamente attraverso la lettura di tale dato oggettivo.
I basidiomi espressi dalle Russule, sono forniti di una loro impostazione morfologica assai semplice e del tutto riconoscibilissima, tale da potersi perfino definire monotona (risulta infatti facilissimo, anche per i principianti, individuare una Russula in quanto tale).
Si tratta di funghi con carposomi costituiti esclusivamente di due componenti specifiche quali, il cappello che ospita la parte fertile e quindi riproduttiva, ed il gambo, struttura sterile a sostegno del cappello stesso. Vediamoli più attentamente:
(1) il cappello o pileo: questo elemento strutturale può assumere, in senso generico, due distinte forme:
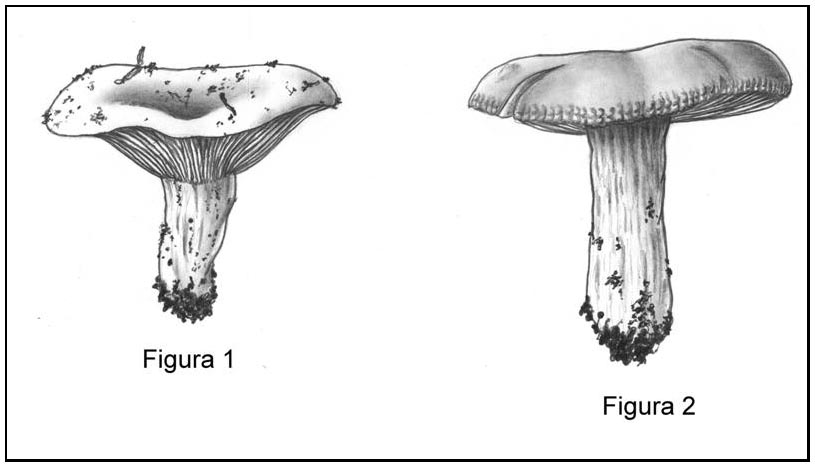
- infundibuliforme (ovvero a guisa di imbuto), configurazione comunque raggiunta a completa maturità, come nel caso di Russula chloroides (Cfr. Figura 1);
- piano convessa e appena depressa al centro, nell’insieme richiamante l’aspetto di un piattino per tazzine (Cfr. Figura 2) o al più di una scodella, il tutto potendo culminare in vecchiaia, nella la classica conformazione a cratere (sagoma raggiunta da un buon numero di specie). (fig. 1 e 2 - cliccare sull'immagine per ingrandire)Il margine della struttura fin qui menzionata, in entrambi i casi, è più o meno regolare. (fig. 1 e 2 - cliccare sull'immagine per ingrandire)Il margine della struttura fin qui menzionata, in entrambi i casi, è più o meno regolare.
La superficie del cappello risulta, sempre ed in ogni caso, rivestita di una pellicola (cuticola) pigmentata, talvolta in modo clamorosamente variopinto.
Nella pagina inferiore del cappello stesso, trova alloggio l’imenoforo, classicamente formato da lamelle (o lame);
(2) il gambo, conosciuto anche come stipite o piede, ha la funzione, come accennato, di supportare la struttura che lo sormonta; è concentrico al cappello ed ha forma grossolanamente cilindrica.
Parlare di dimensioni minime e massime raggiungibili dagli sporofori appartenenti alle specie che fanno capo a questo Genere, risulta alquanto aleatorio e vedremo perché. Comunque è pur vero che a fronte di specie geneticamente use ad esprimere fruttificazioni di dimensioni cospicue, ovvero pari o oltrepassanti i 20 cm di diametro relativi al cappello (comeRussula chloroides, R. nigricans e R. olivacea per citarne i casi più eclatanti), ce ne sono altre di modestissima statura, parliamo di appena 2-4 cm sempre inerenti alla larghezza pileica stessa (ad esempio R. monspeliensis, R. subazureaper chiamare in causa specie presenti in ambito mediterraneo).
Accennavo prima all’aleatorietà concernente il dato dimensionale pertinente a questi funghi, in special modo riferito alle loro variabilità infraspecifiche. E’ un dato di fatto assodato, che specie di complessione media possono differenziare basidiomi dalla stazza inusuale, rispetto alle misure “standard” che comunemente ci è dato osservare.
Ciò dipende, certamente in parte, da fattori per lo più estrinseci, come particolari condizioni pedoclimatiche, nelle quali il micelio si trova a svolgere le proprie funzioni vitali.
Nota è infatti la tendenza al gigantismo dei corpi fruttiferi di molte specie micetiche in ambiente mediterraneo (specialmente in Sicilia), per i quali le dimensioni citate dai libri, contano assai poco.
Scaturenti senz’altro da un’elevata temperatura media giornaliera (escursioni termiche quantitativamente ridotte) e da contemporanei tassi di umidità talvolta consistenti. Concause queste, che di rado coincidono nel medesimo periodo, in special modo nelle zone che frequento assiduamente (estremo ovest della Sicilia), spesso esposte a lunghissimi periodi di aridità. Tuttavia in stagioni particolarmente fortunate, dal punto di vista climatologico, consentono lo sviluppo di sporofori a dir poco eccezionali.
Ho infatti avuto modo di constatare personalmente, quadri dimensionali “titanici” (naturalmente riferiti alla specie) in parecchi basidiomi di R. melliolens, che hanno superato tranquillamente i 20 centimetri di diametro, lo stesso in R. cyanoxantha, perfino in R. fragilis, i quali cappelli si spingevano oltre i 12-13 cm di diametro (dimensioni gigantesche per quest’ultima entità specifica), per non parlare diR. chloroides prossime ai 30 cm.
Tutto ciò, non in più che comprensibili casi sporadici, dovuti alla genesi di fioriture monocarpiche (costituite cioè da un solo “fungo” per un intero micelio), ma altresì regolarmente anche in fruttificazioni gregarie (ossia di sporofori della stessa specie in stretto contatto gli uni con gli altri).
Altresì, condizioni pedoclimatiche poco favorevoli, oltre che in generale ad inibire lo stimolo riproduttivo nel micelio, la differenziazione di sporofori, laddove ciò dovesse verificarsi, non solo avverrebbe in forma numericamente ridotta, ma le proporzioni di questi ne verrebbero drasticamente alterate, tali da dare luogo ad una sorta di nanismo dettato dalle particolari contingenze esogene.
Mi spiego meglio, ho riscontrato che R. persicina (specie rintracciabile ovunque, tanto da risultare “infestante”), in stagioni siccitose non solo riduce al minimo la generazione sporoforale, ma quei pochi basidiomi che i talli miceliari riescono ad estrinsecare, sono di taglia così esigua che a malapena riescono a raggiungere i 3 cm di diametro del cappello ed un‘altezza complessiva di appena 4/5 cm.
Cosa che capita anche alla già citata R. fragilis.
Comunque, una propensione fisiologica (quindi non casuale) al “nanismo”, è strettamente correlata all‘associazione di Russule con taluni partners vegetali.
È ben conosciuta infatti l’esiguità dei basidiomi delle specie aggregate, ad esempio, esclusivamente a Cistus sp. pl.
Strutturazione genetica evoluta dal contraente fungino, certamente in funzione della limitata prestanza del partner fotobionte (in pratica il vegetale sociato al micelio), con conseguente interscambio di nutrienti quantitativamente ristretto.
Una simile causalità biologica, ha di certo Influito sulle necessità vegetative miceliari, sfociate in adattamenti tali da coinvolgere conseguentemente le proprie strutture riproduttive, assestandone le proporzioni (almeno in tali casi) verso standard dimensionali ridotti al minimo indispensabile.
Il Genere Russula in Europa, nel suo complesso, non manifesta basidiomi a crescita fascicolata, cespitosa o conglobata, ma fruttificazioni isolate o gregarie.
Tuttavia sviluppi sporoforali anomali (embrioni di corpi fruttiferi posti talmente vicini gli uni agli altri, da intrecciare e fondere le loro ife durante il progredire della crescita) sono cagione di carposomi concresciuti, tali da essere uniti per la base del gambo o per l’orlo dei cappelli (o per entrambi).d. Accenni sull’Ontogenesi delle Russule
Si intende per “ontogenesi”, l’insieme delle fasi che si succedono durante il corso dello sviluppo di un organismo vivente. Nel caso specifico dei funghi, con tale termine si tende a definire i vari stadi di accrescimento dei carposomi.
Per quel che ho potuto personalmente dedurre dalla bibliografia specialistica inerente al Genere Russula, nella flora europea, i basidiomi presentano uno sviluppo gimnocarpo, ovvero la formazione dell’imenio avviene allo scoperto, senza la protezione di alcun tegumento protettivo (velo di qualsiasi natura).
Tuttavia, alcune Russule del vecchio Continente, manifestano una angiocarpia primaria palese (ossia presenza di struttura velare a protezione dello sporoforo, o meglio di un velo generale).
Più sprecisamente, sarebbe più corretto definire queste entità specifiche monoveloangiocarpiche, dato che in esse è rilevabile la sola presenza del velo primario.
Quattro sono le specie dotate della suddetta struttura velare, che in verità è comunque assolutamente fugace.
Nella fattispecie, queste sono: Russula insignis (taxon assai diffuso anche in area mediterranea), Russula messapica (specie di recentissima classificazione, la cui presenza in Italia è segnalata in massima parte nel Salento), Russula ochroleuca (specie reperibile nelle regioni settentrionali, orbitante nell’arco alpino) ed infine Russula viscida (rintracciabile nel centro Italia in associazione con faggi).
I residui velari cui si è fatta menzione, compaiono in tracce sotto forma di incrostazioni giallastre ben evidenti alla base del gambo, in qualche caso mescolate alla presenza di filamenti araneosi, sia nel tratto basale dello stipite, che al margine del cappello (Russula insignis).
La presenza di ife velari in tracce, è sagnalata dall’uso del KOH (idrato di potassio in soluzione al 20-30%). Laddove, una goccia di tale macroreagente, posta al contatto di queste (come detto in precedenza, specialmente alla base del gambo), infiamma il punto di contatto, di una tinta rossa fuoco veramente notevole.
Tuttavia, (si tratta di mie speculazioni e pertanto confutabilissime), sarebbe lecito chiedersi se anziché essere di fronte ad un apparente sviluppo gimnocarpico, non ci trovassimo invece al cospetto di una gimnangiocarpia (A.F.M. Reijnders, 1963 sullo sviluppo e disposizione dei tegumenti velari), ove la presenza di veli protettivi sia così primitivamente obliterata, da mimare perfettamente un quadro ontogenetico riferibile alla pura gimnocarpia.e. Elementi di istologia nel Genere Russula
I basidiomi delle Russule sono caratterizzati da una trama omogenea (la trama è ciò che viene comunemente definita la carne del fungo), ossia il pileo e lo stipite hanno struttura ifale confluente, in modo tale che dal gambo, essa si continua allo stesso modo, nel cappello senza interruzione di sorta.
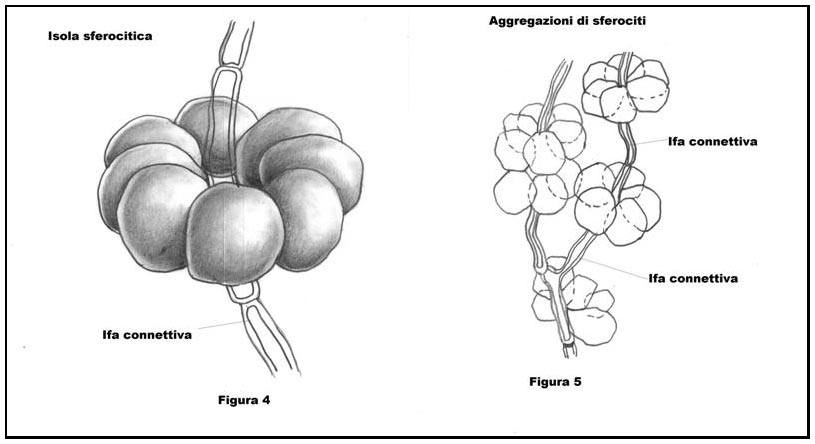
Nel Genere Russula, tale struttura ifale ha una peculiarità (condivisa per altro con l’affine Genere Lactarius), ossia è costituita da sferociti (detti anche sferocisti) o meglio da isole sferocitiche (vedremo qui appresso il significato di tali locuzioni. Cfr. Figura 4: disegno schematico di un’isola sferocitica) che nel loro complesso danno origine ad uno pseudotessuto, cioè un ammasso ifale che ha l’aspetto di un tessuto senza tuttavia esserlo, ove per tessuto è da intendersi una compagine cellulare a divisione mitotica tridimensionale, avente funzione omogenea (per maggiori ragguagli Cfr. Figura 3: Schematizzazione della struttura interna relativa ad uno sporoforo “ideale” di Russula).
Per rendere bene l’idea di quanto appena detto, con un paragone banale, ma calzante, basti pensare al volgarissimo polistirolo espanso (materia plastica leggerissima costituita da agglomerati di piccole sfere). Ebbene la “carne” delle Russule ha una struttura che ricorda questa sostanza sintetica.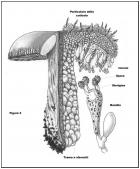
(fig. 3 - cliccare sull'immagine per ingrandire)Ciò giustifica un’altra peculiarità posseduta dalle Russule: la carne “cassante”, ossia che si frattura nettamente (in modo analogo a come si romperebbe un bastoncino di gesso).
Simile caratteristica è dovuta alla particolare conformazione della trama che, ho detto in precedenza, è a struttura sferocitica, la quale sottoposta ad un qualsiasi tipo di flessione, specialmente sul gambo, provoca lo scollamento reciproco di queste cellule grossolanamente sferiche, causandone la rottura in maniera netta.
Per semplificare ulteriormente, basta fare la prova con un listello di polistirolo, si osserverà immediatamente la frattura dello stesso tramite la separazione netta delle “palline” contigue che ne compongono la tessitura, spezzandolo in modo “preciso” e senza sfilacciature.
Come detto in precedenza, gli pseudotessuti della trama nelle Russule sono formate da isole sferocitiche o nidi di sferociti o ancora rosette di sferociti (termini che hanno lo stesso significato), che consistono in aggregazioni di sferocisti, raggruppati i genere attorno ad un ifa connettiva allantoide (Citazione da H. Romagnesi), ove per ifa connettiva allantoide si intende una struttura ifale filamentosa formata da una sequenza di cellule ifali, restringentesi alle estremità, atta al trasporto di sostanze nutritive (Cfr. Figura 4 e 5: Schematizzazione di isole sferocitiche aggregate attorno ad ife connettive).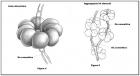 (fig. 4 e 5 - cliccare sull'immagine per ingrandirla) (fig. 4 e 5 - cliccare sull'immagine per ingrandirla)
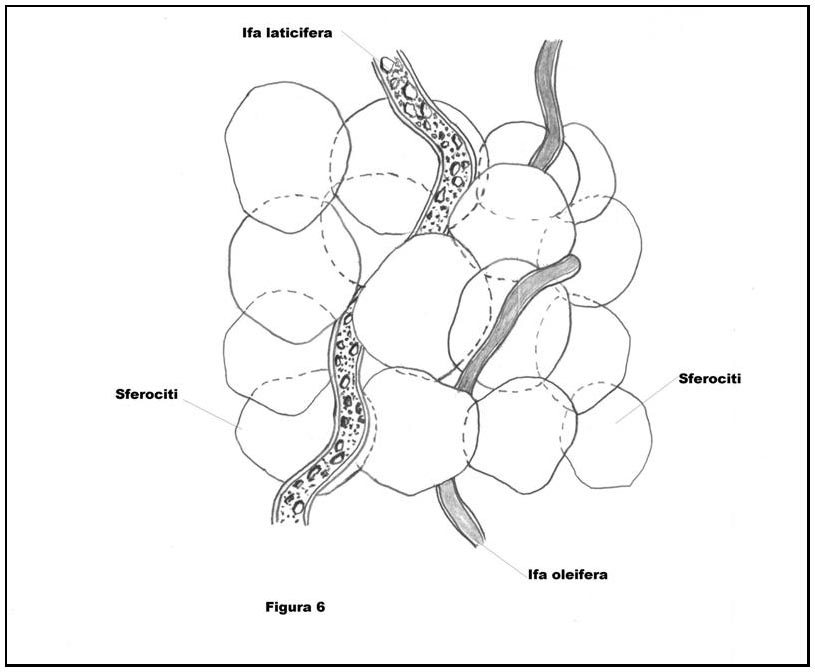
All’interno della trama delle Russule compaiono anche ife escretrici (o ife vascolari), ovvero ife laticifere ed ife oleifere (Cfr. Figura 6: Schematizzazione di ife escretrici presenti nella trama formata da sferociti). 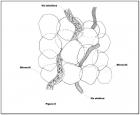
(fig. 6 - cliccare sull'immagine per ingrandirla)
Le prime racchiudono al proprio interno latice, le seconde invece “contengono sostanze generalmente resinose, associate con un sapore acre del carposoma..;” (cito testualmente la Sig.raMaria Teresa Basso dal Volume di cui è autrice “Manuale di microscopia dei funghi”, pag. 109).
E’ da rilevare, per altro come già accennato, che le Russule pur essendo corredate di ife laticifere, come accade nei Lattari, tuttavia non secernono latice né alla frattura né tanto meno all’incisione.
Nelle ife costituenti la struttura dei carposomi, non si rinviene traccia di giunti a fibbia, per cui le specie afferenti a questo Genere ne sono assolutamente prive (ricordo per chiarezza che i giunti a fibbia sono le vestigia delle divisioni mitotiche delle cellule ifali che sequenziano le successive).
La composizione istologica degli sporofori inclusi in questo genere, viene definita di tipo eteromero (ovvero pseudotessuti composti da sferociti misti ad ife filamentose, comprendenti anche le escretrici).
Henri Romagnesi (il padre della Russulologia moderna), notò che più gli sferociti risultano essere di dimensioni ridotte e quindi maggiormente appressati, più la “carne” del carposoma acquisisce durezza e compattezza, per converso sferociti di grandi dimensioni tendono a rammollire e rendere fragile la carne stessa, si vedano gli esempi estremi di Russula lepida (specie nota proprio per la rigidità e la durezza della carne) e Russula fragilis (talmente delicata da sbriciolarsi letteralmente fra le dita se manipolata rudemente).f. Note sulla fenologia
Dal punto di vista fenologico, le Russule, hanno epoche di fruttificazione diversificate. Infatti a fronte di specie che hanno ritmi produttivi tendenzialmente primaverili, moltissime specie invece assolvono a tale incombenza biologica in estate ed in autunno, altre ancora in inverno (chiaramente laddove le temperature medie lo consentano, non scendendo al di sotto di certi parametri).
A mio modestissimo avviso, dettagliare un elenco di specie, in una statistica generalizzata, che contempli la precisa epoca di riproduzione e quindi di differenziazione degli sporofori, di ciascuna di queste, sarebbe un’impresa alquanto lacunosa, per non dire azzardata, e di conseguenza non potrebbe rivestirsi di alcun valore significativo.
A meno che, tale statistica, non fosse elaborata in connessione ad un luogo preciso, analizzato per un’ampiezza temporale distribuita lungo il corso di diversi anni. Solo allora, se ne potrebbero estrapolare dei dati nel merito, con la velleità che questi siano sufficientemente attendibili.
Mi spiego meglio:
è noto che specie quali Russula vesca, Russula cyanoxantha,Russula rubroalba, Russula virescens, Russula romellii,Russula risigallina etc. siano specie a fruttificazione primaverile.
Bene: ho avuto modo di constatare più volte di persona, quanto appena asserito, verificandolo nei castagneti di Galati Mamertino e nelle splendide cerrete di altre località Nebrodensi, per cui presumo sia così anche in consessi floristici similari presenti in tutto il sud-Italia, sebbene a latitudini più settentrionali.
Tuttavia, sarebbe concettualmente errato, tentare di astrarre da queste semplici osservazioni, regole che abbiano una qualche valenza generale.
Infatti, presto verrebbe destituita di ogni fondamento l’idea di considerare per buone, le epoche riproduttive riferite alle specie testè menzionate e riscontrate nelle fascia delle foreste decidue, pretendendo poi, di sovrapporle al Piano Basale Mediterraneo, dominio delle sclerofille sempreverdi, comprendente realtà ecosistemiche ben lontane dalle precedenti.
Zona climatica estrema quest’ultima, è notoriamente contraddistinta da un lunghissimo periodo di aridità annuale, talvolta con siccità protratte dai mesi primaverili ai mesi strettamente estivi, spesso con prolungamenti fino ai mesi primo autunnali, accompagnate da temperature talora elevatissime (in alcuni casi prossime ai 40° C.).
Per cui ogni possibile tentativo di comparare a tale areale fitoclimatico, le periodicità riproduttive delle suddette specie, rilevate altrove, mostrerebbe la sua più totale inadeguatezza, fallendo miseramente.
Quanto appena detto, sarebbe più acutamente apprezzabile nelle sugherete siciliane (cenosi esposte ad una notevole insolazione del soprassuolo con susseguente stress idrico a carico delle componenti botaniche, spesso drammatico), all’interno delle quali, pur annoverandosi la presenza di molte delle specie elencate pocanzi (Russula virescens, Russula vesca, Russula cyanoxantha, e talvolta anche Russula rubroalba), loro fruttificazioni primaverili, sarebbero da considerarsi dei veri e propri miracoli (comunque eccezionalmente accadenti).
In ultima analisi, la conclusione, in tali ambiti vegetazionali, finisce con l’essere il pressochè costante slittamento dello stimolo riproduttivo, nel più favorevole periodo autunnale (e spesse volte nemmeno in quello).
Si può, non di meno, dare un quadro approssimativo a riguardo della Fenologia inerente al Genere Russula. Mi limiterò però alla sola Sicilia, regione della quale ho una qual certa esperienza, facendo soltanto dei brevi cenni sugli ambienti silvani in cui sia possibile reperire questi miceti e, quando, poterli osservare.
Prima di proseguire però, ci tengo comunque ad evidenziare che, in generale, i funghi, affinché possano originare i propri carposomi, abbisognano necessariamente, di determinate condizioni, quali ad esempio un determinato quantitativo d’acqua nel substrato, opportune temperature nell’ambiente di vegetazione, certamente di una massiccia presenza di sostanze nutritive attingibili dal sostrato vitale (la funzione riproduttiva necessità di un ingente apporto energetico) e, virtualmente, di altri fattori concomitanti misconosciuti se non addirittura ancora assolutamente ignoti alla scienza.
Il tutto, è ancora più complicato per la flora fungina micorizzogena (funghi simbionti o micorrizici), la quale dipende, per la propria sopravvivenza, dai partners botanici.
Infatti, qualsiasi eventuale stato di stress in cui potrebbe venire a trovarsi l’ospite vegetale, finirebbe con il ripercuotersi immancabilmente sul micobionte (simbionte fungino), alterandone le funzioni vitali, tanto da comprimerle al solo minimo vegetativo.
In ultima analisi, mancando l’influenza contemporanea di tali contingenze, la spinta riproduttiva del micelio verrebbe del tutto inibita o quanto meno drasticamente modificata.
Quindi, siffatte situazioni, in realtà più frequenti di quel che si creda, finiscono per fare “saltare” qualsiasi schematizzazione volta ad inquadrare tempi di riproduzione fissi.
Detto questo, fornirò ora, dopo le precedenti disquisizioni, un sommario florilegio delle Russule più note, rintracciabili in ciascuno dei contesti forestali in appresso citati, con un’indicazione di massima relativa ai periodi di attività riproduttiva, concernenti queste specie stesse, calate all’interno delle fitocenosi dove si possono reperire.
Il seguente quadro riassuntivo, investe esclusivamente i sistemi silvani maggiormente presenti sul territorio regionale Siciliano.
- Le sugherete, la macchia mediterranea e i cisteti: vantano una ricchissima presenza di Russule. Assumiamo quale dato certo, il fatto che eventuali fruttificazioni primaverili siano evento raro (non impossibile), e spostiamo il tutto ai mesi autunnali (ferme restando vi siano le opportune condizioni indispensabili).
- Settembre/Ottobre: le prime specie a riprodursi sono sicuramente le xero-termofile, quindi Russula vesca, Russula persicina, Russula werneri, Russula chloroides, Russula grisea, Russula sororia, Russula virescens, Russula amoenicolor ecc.
- Ottobre/Novembre: continuano le specie termofile, quindi Russula seperina, Russula persicina ancora, Russula heterophylla, Russula chloroides, Russula cyanoxantha e forme correlate; poi Russula graveolens e forme correlate
- Novembre/Dicembre: Ancora Russula graveolens e forme correlate, resiste Russula persicina la cui presenza si va facendo sempre più rarefatta, Russula amoenolens, Russula subazurea, compare Russula fragilis, che continua anche fino a Gennaio nella annate poco fredde, dimostrandosi in tali contesti la specie più criofila fra le Russule.
- Le leccete: anche in tali complessi vi è una buona presenza di Russule (ma inferiore alla sughereta).
- Settembre/Ottobre: le prime specie a riprodursi sono Russula vesca, Russula persicina, Russula ilicis, Russula chloroides, Russula insignis, Russula praetervisa, Russula odorata, Russula amoenicolor ecc.
- Ottobre/Novembre: Russula sperina, Russula persicina ancora, Russula maculata, Russula chloroides, Russula cyanoxantha e forme correlate; anche qui Russula graveolens e forme correlate; Russula pelargonia
- Novembre/Dicembre: Ancora Russula graveolens e forme correlate e Russula pelargonia.
- Le pinete di rimboschimento: Nei suddetti ambienti, dove in pratica si è proceduto alla riforestazione, utilizzando l’onnipresente Pinus halepensis misto in qualche caso a Pinus pinea (pino domestico), scarsa è la presenza di Russule, vi troviamo infatti la stessa Russula chloroides nel periodo primo autunnale, fino al mese di Novembre, e poi Russula torulosa e forme relazionate, essendo le Russule stesse in simili espressioni arboree, sostituite dal Genere Lactarius (Lactarius deliciosus, Lactarius sanguifluus e la sua varietà vinosus), salvo i casi in cui ai pini non si siano consorziati i cisti o l’erica, nell’ipotesi, il quadro muterebbe, avendo tali essenze un discreto peso nell’implementare un proprio corteggio fungino.
- Il castagneto e il querceto deciduo: Laddove siano insediate le essenze arboree decidue, delle quali il Castagno (Castanea sativa) è una delle rappresentati più note, o il querceto caducifoglio, nel nostro caso formato dalla dominanza del Cerro (Quercus cerris) la presenza delle Russule presenta una buona varietà di specie, tuttavia l’arco temporale in cui può avvenire la riproduzione, si sposta nei mesi primaverili oppure tardo estivi e primo autunnali, andando poi le piante in quiescienza invernale con l’interruzione momentanea, o quanto meno, ridotta al minimo, del rapporto ectotrofico con le specie micorrizogene satelliti.
- Maggio: le prime specie a riprodursi in tali ambiti sono Russula vesca, Russula virescens, Russula rubroalba, Russula romellii, Russula cyanoxantha e forme correlate, Russula lepida, Russula melliolens, Russula albonigra, Russula heterophylla, Russula risigallina, tutte specie tipiche, che si possono incontrare anche a fine Agosto e Settembre.
- La faggeta: In Sicilia il Faggio (Fagus sylvatica), costituisce in generale il livello altitudinale estremo della vegetazione arborea, si tratta anche stavolta di un’essenza decidua, seguita da una discreta presenza di Russule, eminentemente a riproduzione primaverile ed estiva, in qualche caso prolungantesi nei mesi di settembre e Ottobre (annate calde). Vi troviamo Russula albonigra, Russula vesca, Russula heterophylla, Russula virescens, Russula romellii, Russula cyanoxantha, Russula rubroalba, Russula amoenicolor, Russula foetens, Russula melliolens, Russula faginea, Russula olivacea, Russula risigallina ecc.
Ripeto, questi riferimenti, sia inerenti agli ambienti naturali citati (o artificiali) che ai periodi di comparsa ed alle specie per ciascuno menzionate, sono assolutamente semplificati e non hanno nessuna altro scopo che essere mere indicazioni. g. Cenni di Ecologia
L’intera micoflora Europea, inclusa nel Genere Russula, comprende funghi univocamente simbionti, non esistendo fra di esse, entità saprobe o parassite.
Ciò significa che le Russule sono in grado di svolgere le proprie funzioni vitali solo attraverso legame trofico attivato dal mutuo scambio di sostanze nutritive, con essenze vegetali arboree o arbustive.
L’interscambio fra partner fungino e ospite fotobionte avviene, dapprima attraverso l’infezione da parte del micelio (che è in realtà il vero organismo “fungo”) ai “danni” dell’apparato radicale arboreo o arbustivo quale che esso sia.
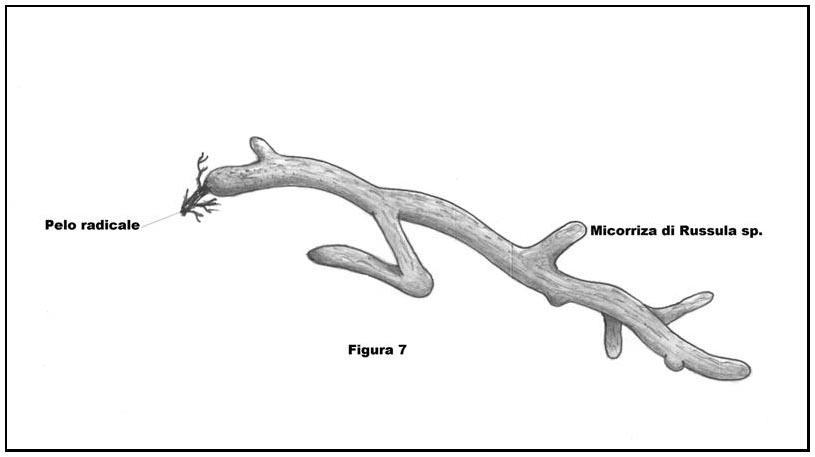
All’avvenuta stabilizzazione dello status definitivo di contraenti la simbiosi, il micelio fungino genera attorno ai peli radicali una struttura composta da ife compattate, dalle sembianze di un manicotto, da cui si dipartono diverse ramificazioni, chiamato micorriza (Cfr. Figura 7: Rappresentazione di una Micorriza in Russula sp). (fig. 7 - cliccare sull'immagine per ingrandire) (fig. 7 - cliccare sull'immagine per ingrandire)
Da qui il nome di funghi micorrizici, quindi la micorriza stessa non è che l’elemento pseudotissutale attraverso cui avvengono gli scambi nutrizionali già menzionati.
Le Russule differenziano micorrize ectotrofiche. La micorriza ectotrofica è costituita da un mantello esterno chiamato micoclena cui segue, verso l’interno, uno strato ifale conosciuto come reticolo di Hartig, le cui ife si interfacciano alle cellule radicali senza tuttavia penetrarne la parete invadendone il lume interno.
Si operano a quel livello, gli scambi di sostanze nutritive (acqua, sali minerali, auxine ecc. dal fungo alla pianta. Idrati di carbonio dalla pianta al fungo).
Dal punto di vista delle essenze con cui contrarre simbiosi mutualistica, fra le Russule si trovano specie ubiquitarie, ovvero reperibili nei più disparati ambienti boschivi. Tipico è il caso di Russula cyanoxantha i cui areali di presenza abbracciano le foreste centroeuropee, fino a spingersi nei caldi boschi termo-xerofili mediterranei, passando per tutti i piani e gli orizzonti vegetazionali intermedi (ad esclusione... “della flora artico-alpina” – M. Sarnari: Monografia illustrata del Genere Russula in Europa. Tomo 1°. pag. 236).
Altre specie sono a diffusione esclusivamente settentrionale, come ad esempio Russula mustelina, comune nelle coniferete di montagna in compagnia di Picea abies (il famoso peccio o abete rosso), di Larix decidua (larice) e Pinus cembra (cembro). Anche le Abetine, ossia foreste con dominanza di Abies alba (abete bianco) contano su un discreto numero di russule nelle loro compagini micofloristiche.
Altri pini, che hanno un discreto corteggio di Russule sono Pinus sylvestris (pino silvestre), Pinus nigra (pino nero) ed essenze più termofile, prettamente mediterranee quali Pinus pinaster (pino marittimo), Pinus pinea (pino domestico) e Pinus halepensis (pino d’Aleppo)
Altre ancora sono tipiche dei comparti forestali decidui e le troviamo in selve dove domini Fagus sylvatica (faggio), oppure a seguito di Betula pendula (betulla).
Anche nei mesofili querceti decidui, puri o misti ad altre latifoglie la presenza di Russule è di assoluta rilevanza. Così come accade per le foreste di Quercus cerris (cerro), a cui si mescola, nelle quote più basse di questa fascia vegetazionale, una essenza semidecidua quale Quercus pubescens (roverella).
E ancora, sono presenti negli igrofili e ormai quasi estinti boschi planiziari, che un tempo erano patrimonio della Pianura Padana, ridotti ormai a sparuti paesaggi vegetazionali relittuali, in cui spiccano Quercus petrea (Rovere) e Quercus robur (farnia).
Si rintracciano Russule anche nei misti in cui vi sia la presenza di Populus tremula (pioppo). Oppure in formazioni con dominanza di Ostrya carpinifolia (carpino nero). Laddove Castanea sativa (castagno), offra lo spettacolo delle sue suggestive selve, troviamo un corteggio di russule assai significativo.
Altre specie ancora, sotto il profilo del partner fotobionte, possono avere preferenze specie specifiche molto spiccate, apparendo simbionti obbligati a legame esclusivo con determinate essenze arboree, quali ad esempio Russula alnetorum e Russula pumila, ectomicorrizogene sia di Alnus glutinosa (ontano comune) che Alnus incana (ontano bianco).
Arrivando nei caldissimi boschi mediterranei, regno incontrastato delle foreste sempreverdi, nelle quali spiccano Quercus ilicis (leccio) e Quercus suber (sughera), troviamo il paradiso del cosmo russulologico (in ispecial modo nelle sugherete), con la presenza di numerosissime specie (passibili di aumentare, vista la non ancora definitivamente investigata micoflora mediterranea).
Chiudo questa carrellata di ecosistemi, accennando infine ai due estremi opposti, che chiudono l’Italia a nord e a sud: ovvero le Microselve Alpine, allogate al di là del limite della vegetazione arborea, ben oltre i 2.000 mt di quota, fino ai 2.700 mt s.l.m. (limite vicino alle nevi perenni) e i torridi Cisteti mediterranei prossimi alla fascia litoranea. Costituiti da formazioni monospecifiche o consorziate ad altre entità fitocenotiche di gariga.
Entrambe questi sistemi ecologici presentano condizioni assai difficili e, come già detto estreme. Sono questi, mondi a parte, che contano su una flora micorizogena, includente anche una ben definita quota di Russule, grandemente selezionata e meravigliosamente adattata a sopravvivervi.
Tipiche delle Microselve Alpine sono ad esempio Russula pascua, Russula chamitae, Russula dryadicola ecc.
Russula monspeliensis e Russula cistoadelpha, ecc. sono invece caratteristiche delle roventi cistaie.
I caratteri edafici dei suoli, relativamente alle proprietà chimiche da essi possedute, sembrano avere una certa importanza per le cenosi fito-micologiche
Infatti, come ormai noto, il pH dei terreni gioca un ruolo di primo piano, sulla presenza di talune popolazioni botaniche rispetto ad altre, in un determinato biotopo, influenzandone anche la presenza delle specie fungine.
Per pH, in chimica acronimo che significa potenza d’idrogeno (simbolo H), si intende la basicità o l’acidità di una soluzione acquosa.
Essa è misurabile attraverso un indice di riferimento servendosi della famosa cartina al tornasole o di un particolare strumento chiamato potenziometro.
Il valore 7 denuncia che la soluzione è neutra (cioè che non è acida nè tantomeno basica). Valori inferiori a 7 indicano livelli di acidità via via sempre più crescenti al decrescere del numero in indice. Altresì valori superiori al 7 sottintendono la basicità della soluzione che presenta livelli maggiori al crescere del numero nell’indice stesso
Gli organismi viventi che hanno bisogno di vivere in ambiente acido si chiamano acidofili.
Quelli che vivono in ambienti basici si chiamano invece basofili.
Il discorso sull’acidità o la basicità del mezzo non è valida solo e soltanto per le soluzioni acquose, ma anche per la definizione dei suoli, esistendo infatti terreni a pH acido, neutro o basico, in dipendenza della struttura pedologica che questi presentano.
Come ben si conosce, ad esempio, suoli ricchi di sabbie e quindi silicei, sono per loro natura neutro-acidi o acidi, mentre suoli ricchi di carbonati di calcio, presentano un discreto livello di basicità.
Le piante, sono organismi viventi strettamente dipendenti dai valori di pH presenti nella composizione chimica dei terreni di vegetazione, per cui alcune di esse sono talmente adattate a vivere in suoli acidi o basici oppure neutro-acidi o neutro-basici, tanto che difficilmente sopravvivrebbero al mutare di tali fattori.
Anche i funghi rispondono a queste regole ecologiche e le Russule non sono da meno.
Per cui a fronte di specie a propensione acidofila, ve ne sono altre a tendenza basofila, altre ancora assolutamente indifferenti alla natura chimica del substrato.
Credo comunque, che complessivamente, le Russule siano miceti tendenzialmente neutri dal punto di vista del gradimento chimico dei suoli, e dotati di una resilienza tale, da permettere loro di adattarsi alle fluttuazione dei pH ove intercettare gli ospiti vegetali cui aggregarsi.
2. COME PROCEDERE NELLO STUDIO DEI FUNGHI Concluso il discorso generale oggetto del precedente paragrafo, mi permetto una necessaria digressione che ci consentirà di procedere in maniera organica nell’approfondimento analitico delle Russule (ma applicabile anche per tutti gli altri taxa fungini).
Lo studio di una qualsiasi specie micetica, viene ancora in larga parte effettuata utilizzando per lo scopo, gli sporofori o basidiomi (nel caso di basidiomiceti) o anche corpi fruttiferi, insomma la parte riproduttiva e allo stesso tempo visibile, per periodi annuali limitatissimi del micelio (generalmente in primavera e autunno), quello che noi erroneamente identifichiamo come fungo, atteso che il vero fungo sia invece il micelio o tallo vegetativo.
L’esame approfondito e meticoloso dei corpi fruttiferi serve essenzialmente per i seguenti motivi.
a. Classificazione, operazione da effettuare nel caso in cui ci si imbatta eventualmente in una specie sconosciuta fino ad allora, al fine di conferirle una identità e collocarla all’interno di uno schema tassonomico preciso. Ciò comporta il rispetto di una procedura contenuta nel codice di nomenclatura botanica internazionale. Facciamo un esempio pratico: trovo in una lecceta una entità “strana”, che so comunque già appartenere al Genere Russula. Dallo studio compiuto, mi rendo conto con certezza, che essa non rientra fra le specie finora conosciute appartenenti a tale Genere. Quindi, seguendo le norme contemplate nel codice suddetto, le attribuisco un epiteto (ossia un nome) specifico scegliendone uno non ancora esistente per quel Genere, che poniamo il caso sia xanthopurpurea(prendendo spunto dalla colorazione tipica del cappello). Ne compilo la diagnosi in latino completa di tutti i caratteri, ivi compresa la microscopia, allego la foto e l'exiccatum e deposito il tutto nelle adeguate sedi. Ecco classificata una nuova entità, Russula xanthopurpurea, appunto.
b. Determinazione, approfondimento condotto invece su sporofori appartenenti ad una entità specifica già presumibilmente nota alla scienza, ma tuttavia ignota al ritrovatore, il quale procede allo studio della raccolta per identificarla, utilizzando metodi e materiali atti allo scopo.
c. Riconoscimento, altro non è che l’immediata identificazione dello sporoforo sul campo, perché, dietro nostra esperienza personale, già conosciamo bene la specie reperita. Lo studio dei funghi si avvale di diversi tipi di indagine, condotti come già detto, direttamente sugli sporofori (o carposomi, o basidiomi nel caso di basidiomiceti), che nel loro insieme ci condurranno all’esito di dare un nome alla nostra raccolta. Gli esami da condurre sono i seguenti, il tutto naturalmente deve essere corredato da apposita iconografia del reperto prelevato (in pratica una bella foto che ritragga il “fungo” in più stadi di crescita mettendone in evidenza tutti gli elementi strutturali):
(1) Caratteri ecologici: rilevamento dell’habitat di crescita con indicazione delle essenze vegetali trovate sul posto (questo implica la conoscenza delle fondamentali essenze arboree e arbustive degli ambienti naturali che frequentiamo);
(2) Caratteri macroscopici:
- forme e dimensioni, pigmentazioni delle strutture che compongono gli sporofori, qualità della trama o carne ivi compresi eventuali viraggi;
- qualifica del colore della sporata (particolarmente importante per il Genere Russula).
(3) caratteri organolettici:
- odore emanato dagli sporofori;
- sapore posseduto da questi.
(4) Caratteri chimici:
- macrochimismo degli sporofori utilizzando allo scopo particolari reagenti detti macrochimici;
- microchimismo degli sporofori utilizzando allo scopo particolari reagenti per le strutture istologiche da rilevare al microscopio ottico.
(5) Caratteri microscopici:
icludendo naturalmente il microchimismo, forma, dimensioni e ornamentazioni delle spore, cuticola pileica (pileipellis), struttura delle ife, pileocistidi (o altro) e imenio ossia basidi e cistidi.
Fatti tutti quanti questi passaggi, si passa alla redazione della scheda, mettendo su carta la descrizione del nostro ritrovamento (dopo aver avuto l’accortezza di essiccare gli sporofori reperiti previa loro riduzione in fette sottili e conservando il tutto. ivi compresa la sporata ottenuta, dentro una bustina di plastica).
A questo punto, ricorrendo all’ausilio di una flora analitica, ovvero un testo basato sull’uso di chiavi dicotomiche, cerchiamo di risalire al Genere e di arrivare quanto più possibile vicino alla specie che abbiamo raccolto. Quindi si passa alla consultazione delle opere monografiche, ovvero testi specialistici dedicati ad un solo Genere (Amanita, Entoloma, Russula ecc.).
Incrociando le nostre ricerche con i risultati desunti da altri testi ancora dovremmo ottenere la definitiva determinazione della specie studiata.
Nei casi più ostici e dubbi, laddove avessimo ancora delle perplessità, malgrado lo studio sui testi, si passa a richiedere un exiccatum (porzione disseccata di un qualsiasi sporoforo) della specie che crediamo di aver individuato. Si procede allora, al confronto diretto tra il nostro materiale, relativamente alla microscopia che ne abbiamo ottenuto, paragonandolo (dopo opportuno prelievo) al reperto secco ricevuto. Se dall’esame condotto, si notano evidentissime affinità nei quadri istologici e citologici, tra specie reperita ed exiccatum stesso, abbiamo determinato con sicurezza la specie presunta.
Per studiare con successo le Russule occorrono:
- una buona macchina fotografica digitale, ormai alla portata di tutti, quanto meno munita di zoom;
- un taccuino su cui annotare la vegetazione presente sul luogo di reperimento, l’odore ed il sapore testati al momento della raccolta, nonché eventualmente il colore della cuticola;
- reagenti macrochimici (vedremo nella 2ª parte di questo contributo quali essi siano);
- reagenti microchimici e coloranti di contrasto (anche in questo caso si darà descrizione del materiale nella 2ª parte di questo contributo);
- un buon microscopio ottico, preferibilmente binoculare, munito di oculare micrometrico per le misurazioni degli elementi istologici e corredato di un obiettivo 100x ad immersione;
- testi appositi per lo studio di questi funghi (se ne darà un elenco nella 2ª parte di questo contributo).
Concludo qui il segmento iniziale di questo lavoro, spero di non essere stato troppo pesante, e di avere reso chiaro ai lettori alcuni aspetti generali su questo affascinate argomento. Mi scuso, qualora eventualmente non fossi risuscito nell’intento di rendere cristallina l’esposizione, tuttavia mi sono sforzato di farlo nel migliore dei modi. Se errori concettuali dovessero essere presenti in questo scritto, ne sono l’unico responsabile.
Ci rivedremo per la seconda parte ove concluderemo il discorso qui iniziato.Grazie per avermi seguito lungo tutte queste pagine.
Nino Mannina
3. BIBLIOGRAFIA DI RIFERIMENTO
- I Boschi di Sicilia. Edizioni Arbor 1992
- Cappelletti Carlo: Botanica Voll. 1 e 2. Utet Torino. 1975
- Martin Elizabeth: Conoscere gli alberi. Edizioni Euroclub 1980
- Paci Marco: Ecologia Forestale. Edizioni Edagricole 1997
- Basso Maria Teresa. Manuale di microscopia dei funghi. Libreria Mykoflora 2005
- Rambelli A. e Pasqueletti M.: Nuovi fondamenti di Micologia. Jaca Book 1996
- Galli Roberto: Le Russule. Edinatura 1996
- Romagnesi Henri: Les Russules d’Europe et d’Afrique du Nord. A.R.G. Gantner Verelag K.-G. Vaduz Reprint 1996
- Sarnari Mauro: Monografia illustrata dl Genere Russula in Europa Tomo 1. A.M.B. Fondazione Studi Micologici 1998
- Sarnari Mauro: Monografia illustrata dl Genere Russula in Europa Tomo 2. A.M.B. Fondazione Studi Micologici 2005
Copyright:
Testo e disegni inseriti in questo approfondimento sono riservati e di esclusiva proprietà dell'autore: Nino Mannina.
Ringraziamenti: L'autore ringrazia l'amico Nino Giacalone, Presidente del Gruppo Micologico "TONINO POCOROBBA", per aver curato impostazione e inserimento di questo mio scritto nel nostro sito, come sempre in maniera impeccabile ed elegante.
|